Ионные каналы проводят ионы сквозь биологическую мембрану. Заболевания, связанные с ионными каналами. Потенциалзависимые натриевые каналы
Согласно современным представлениям, биологические мембраны образуют наружную оболочку всех животных клеток и формируют многочисленные внутриклеточные органеллы. Наиболее характерным структурным признаком является то, что мембраны всегда образуют замкнутые пространства, и такая микроструктурная организация мембран позволяет им выполнять важнейшие функции.
Строение и функции клеточных мембран.
1. Барьерная функция выражается в том, что мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. При этом мембрана принимает участие в механизмах электрогенеза. К ним относятся механизмы создания потенциала покоя, генерация потенциала действия, механизмы распространения биоэлектрических импульсов по однородной и неоднородной возбудимым структурам.
2. Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»).
3. Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах).
4. Высвобождение нейромедиаторов в синаптических окончаниях.
Современными методами электронной микроскопии была определена толщина клеточных мембран (6-12 нм). Химический анализ показал, что мембраны в основном состоят из липидов и белков, количество которых неодинаково у разных типов клеток. Сложность изучения молекулярных механизмов функционирования клеточных мембран обусловлена тем, что при выделении и очистке клеточных мембран нарушается их нормальное функционирование. В настоящее время можно говорить о нескольких видах моделей клеточной мембраны, среди которых наибольшее распространение получила жидкостно-мозаичная модель.
Согласно этой модели, мембрана представлена бислоем фосфолипидных молекул, ориентированных таким образом, что гидрофобные концы молекул находятся внутри бислоя, а гидрофильные направлены в водную фазу. Такая структура идеально подходит для образования раздела двух фаз: вне- и внутриклеточной.
В фосфолипидном бислое интегрированы глобулярные белки, полярные участки которых образуют гидрофильную поверхность в водной фазе. Эти интегрированные белки выполняют различные функции, в том числе рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул.
Некоторые белковые молекулы свободно диффундируют в плоскости липидного слоя; в обычном состоянии части белковых молекул, выходящие по разные стороны клеточной мембраны, не изменяют своего положения.
Электрические характеристики мембран:
Емкостные свойства в основном определяются фосфолипидным бислоем, который непроницаем для гидратированных ионов и в то же время достаточно тонок (около 5 нм), чтобы обеспечивать эффективное разделение и накопление зарядов и электростатическое взаимодействие катионов и анионов. Кроме того, емкостные свойства клеточных мембран являются одной из причин, определяющих временные характеристики электрических процессов, протекающихщих на клеточных мембранах.
Проводимость (g) - величина, обратная электрическому сопротивлению и равная отношению величины общего трансмембранного тока для данного иона к величине, обусловившей его трансмембранной разности потенциалов.
Через фосфолипидный бислой могут диффундировать различные вещества, причем степень проницаемости (Р), т. е. способность клеточной мембраны пропускать эти вещества, зависит от разности концентраций диффундирующего вещества по обе стороны мембраны, его растворимости в липидах и свойств клеточной мембраны.
Проводимость мембраны является мерой ее ионной проницаемости. Увеличение проводимости свидетельствует об увеличении количества ионов, проходящих через мембрану.
Строение и функции ионных каналов . Ионы Na+, K+, Са2+, Сl- проникают внутрь клетки и выходят наружу через специальные, заполненные жидкостью каналы. Размер каналов довольно мал.
Все ионные каналы подразделяются на следующие группы:
- По избирательности:
a) Селективные, т.е. специфические. Эти каналы проницаемы для строго определенных ионов.
b) Малоселективные, неспецифические, не имеющие определенной ионной избирательности. Их в мембране небольшое количество.
- По характеру пропускаемых ионов:
a) калиевые
b) натриевые
c) кальцевые
d) хлорные
- По скорости инактивации, т.е. закрывания:
a) быстроинактивирующиеся, т.е. быстро переходящие в закрытое состояние. Они обеспечивают быстро нарастающее снижение МП и такое же быстрое восстановление.
b) медленноинактирующиеся. Их открывание вызывает медленное снижение МП и медленное его восстановление.
4. По механизмам открывания:
a) потенциалзависимые, т.е. те которые открываются при определенном уровне потенциала мембраны.
b) хемозависимые, открывающиеся при воздействии на хеморецепторы мембраны клетки физиологически активных веществ (нейромедиаторов, гормонов и т. д).
В настоящее время установлено, что ионные каналы имеют следующее строение:
1.Селективный фильтр, расположенный в устье канала. Он обеспечивает прохождение через канал строго определенных ионов.
2.Активационные ворота, которые открываются при определенном уровне мембранного потенциала или действии соответствующего ФАВ. Активационные ворота потенциалзависимых каналов имеется сенсор, который открывает их на определенном уровне МП.
3.Инактивационные ворота, обеспечивающие закрывание канала и прекращение проведения ионов по каналу на определенном уровне МП.(Рис).
Неспецифические ионные каналы не имеют ворот.
Селективные ионные каналы могут находиться в трех состояниях, которые определяются положением активационных (м) и инактивационных (h) ворот:
1.Закрытом, когда активационные закрыты, а инактивационные открыты.
2.Активированном, и те и другие ворота открыты.
3.Инактивированном, активационные ворота открыты, а инактивационные закрыты
Функции ионных каналов:
1. Калиевый (в покое) – генерация потенциала покоя
2. Натриевый – генерация потенциала действия
3. Кальциевый - генерация медленных действий
4. Калиевый (задержанное выпрямление) – обеспечение реполяризации
5. Калиевый кальций-активируемый – ограничение деполяризации, обусловленной током Са+2
Функцию ионных каналов изучают различными способами. Наиболее распространенным является метод фиксации напряжения, или «voltage-clamp». Сущность метода заключается в том, что с помощью специальных электронных систем в процессе опыта изменяют и фиксируют на определенном уровне мембранный потенциал. При этом измеряют величину ионного тока, протекающего через мембрану. Если разность потенциалов постоянна, то в соответствии с законом Ома величина тока пропорциональна проводимости ионных каналов. В ответ на ступенчатую деполяризацию открываются те или иные каналы, соответствующие ионы входят в клетку по электрохимическому градиенту, т. е. возникает ионный ток, который деполяризует клетку. Это изменение регистрируется с помощью управляющего усилителя и через мембрану пропускается электрический ток, равный по величине, но противоположный по направлению мембранному ионному току. При этом трансмембранная разность потенциалов не изменяется.
Изучение функции отдельных каналов возможно методом локальной фиксации потенциала «path-clamp». Стеклянный микроэлектрод (микропипетка) заполняют солевым раствором, прижимают к поверхности мембраны и создают небольшое разрежение. При этом часть мембраны подсасывается к микроэлектроду. Если в зоне присасывания оказывается ионный канал, то регистрируют активность одиночного канала. Система раздражения и регистрации активности канала мало отличается от системы фиксации напряжения.
Ток через одиночный ионный канал имеет прямоугольную форму и одинаков по амплитуде для каналов различных типов. Длительность пребывания канала в открытом состоянии имеет вероятностный характер, но зависит от величины мембранного потенциала. Суммарный ионный ток определяется вероятностью нахождения в открытом состоянии в каждый конкретный период времени определенного числа каналов.
Наружная часть канала сравнительно доступна для изучения, исследование внутренней части представляет значительные трудности. П. Г. Костюком был разработан метод внутриклеточного диализа, который позволяет изучать функцию входных и выходных структур ионных каналов без применения микроэлектродов. Оказалось, что часть ионного канала, открытая во внеклеточное пространство, по своим функциональным свойствам отличается от части канала, обращенной во внутриклеточную среду.
Именно ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость.
Селективность, или избирательность, канала обеспечивается его особой белковой структурой. Большинство каналов являются электроуправляемыми, т. е. их способность проводить ионы зависит от величины мембранного потенциала. Канал неоднороден по своим функциональным характеристикам, особенно это касается белковых структур, находящихся у входа в канал и у его выхода (так называемые воротные механизмы).
Рассмотрим принцип работы ионных каналов на примере натриевого канала. Полагают, что в состоянии покоя натриевый канал закрыт. При деполяризации клеточной мембраны до определенного уровня происходит открытие m-активационных ворот (активация) и усиление поступления ионов Na+ внутрь клетки. Через несколько миллисекунд после открытия m-ворот происходит закрытие h-ворот, расположенных у выхода натриевых каналов (инактивация). Инактивация развивается в клеточной мембране очень быстро и степень инактивации зависит от величины и времени действия деполяризующего стимула.
При генерации одиночного потенциала действия в толстом нервном волокне изменение концентрации ионов Na+ во внутренней среде составляет всего 1/100000 от внутреннего содержания ионов Na гигантского аксона кальмара.
Кроме натриевых, в клеточных мембранах установлены другие виды каналов, избирательно проницаемых для отдельных ионов: К+, Са2+, причем существуют разновидности каналов для этих ионов.
Ходжкин и Хаксли сформулировали принцип «независимости» каналов, согласно которому потоки натрия и калия через мембрану независимы друг от друга.
Свойство проводимости различных каналов неодинаково. В частности, для калиевых каналов процесс инактивации, как для натриевых каналов, не существует. Имеются особые калиевые каналы, активирующиеся при повышении внутриклеточной концентрации кальция и деполяризации клеточной мембраны. Активация калий-кальцийзависимых каналов ускоряет реполяризацию, тем самым восстанавливая исходное значение потенциала покоя.
Особый интерес представляют кальциевые каналы. Входящий кальциевый ток, как правило, недостаточно велик, чтобы нормально деполяризовать клеточную мембрану. Чаще всего поступающий в клетку кальций выступает в роли «мессенджера», или вторичного посредника. Активация кальциевых каналов обеспечивается деполяризацией клеточной мембраны, например входящим натриевым током.
Процесс инактивации кальциевых каналов достаточно сложен. С одной стороны, повышение внутриклеточной концентрации свободного кальция приводит к инактивации кальциевых каналов. С другой стороны, белки цитоплазмы клеток связывают кальций, что позволяет поддерживать длительное время стабильную величину кальциевого тока, хотя и на низком уровне; при этом натриевый ток полностью подавляется. Кальциевые каналы играют существенную роль в клетках сердца. Электрогенез кардиомиоцитов рассматривается в главе 7. Электрофизиологические характеристики клеточных мембран исследуют с помощью специальных методов.
Краткое описание:
Сазонов В.Ф. Ионные каналы мембраны [Электронный ресурс] // Кинезиолог, 2009-2017: [сайт]. Дата обновления: 31.01.2017..__.201_). _Обзор ионных каналов мембраны. Определение понятия "ионные каналы", их строение, свойства, функции, функциональные состояния, функциональная классификация.
Введение
Ионные каналы (ИК) клеточной мембраны имеют огромное значение для жизни клеток. Они обеспечивают обмен клетки с окружающей средой веществом, энергией и информацией, с них начинаются и ими поддерживаются процессы возбуждения и торможения в нервной системе и мышцах, именно они (вместе и другими молекулярными рецепторами) обеспечивают восприятие клеткой внешних сигналов. С помощью ИК происходит передача в клетку управляющих сигналов из окружающей её среды. Именно ИК обеспечивают синаптическую передачу возбуждения от возбуждённого нейрона на другие клетки. Обобщая, можно сказать, что почти все важнейшие физиологические процессы в организме начинаются с ионных каналов и поддерживаются ими!
Определение понятия
Ионные каналы мембраны - это маленькие белковые трубочки разного диаметра, вставленные в клеточную мембрану, через которые внутрь клетки или наружу могут перемещаться ионы. Перемещение ионов через ионные каналы приводит к изменению концентрации ионов внутри и снаружи клетки, а также к изменению электрического потенциала мембраны. Перемещение в клетку ионов кальция через кальциевые каналы запускает в ней различные внутренние биохимические процессы. Существует множество видов ионных каналов. © 2014-2017 Сазонов В.Ф. © 2014-2016 kineziolog.bodhy.ru..
Ионный канал клеточной мембраны - это отверстие в мембране, обмётанное по краям белковой нитью, через которое через мембрану могут перемещаться ионы. Белковая нить нужна для того, чтобы отверстие не затянулось жировым слоем мембраны. Во многих случаях белковая нить, или каналообразующий белок, обладает функциональной активностью и контролирует пропускную способность канала по отношению к различным ионам. © 2014-2017 Сазонов В.Ф. © 2014-2016 kineziolog.bodhy.ru..
Итак, ионные каналы (ИК) - это сложные трансмембранные белковые структуры, пронизывающие клеточную мембрану поперёк в виде нескольких петель и образующие в мембране сквозное отверстие (пору). Канальные белки состоят из субъединиц, образующих структуру со сложной пространственной конфигурацией, в которой кроме поры обычно имеются дополнительные молекулярные системы: открытия, закрытия, избирательности, инактивации, рецепции и регуляции. ИК могут иметь не один, а несколько участков (сайтов) для связывания с управляющими веществами (лигандами).
Ионные каналы можно рассматривать как транспортный механизм , обеспечивающий перемещение ионов между цитоплазмной клетки и наружной средой.
Кроме ИК в мембране суцществуют и другие транспортные системы для переноса через неё различных веществ (смотрите транспортные механизмы мембраны). Так, перенос веществ может осуществляться специальными транспортными белками , или транслоказами . Транслоказы - это несколько иное понятие, чем ИК. В отличие от мембранных каналов , транслоказы в процессе переноса вещества через мембрану взаимодействуют с ним как с лигандом и при этом претерпевают конформационные изменения . По кинетике перенос веществ с помощью транслоказ в виде облегчённой диффузии напоминает ферментативную реакцию.
Упрощённое определение:
Ионные каналы - это поры (дырочки) в клеточной липидной мембране, которые "обшиты" по краям белковой нитью, чтобы дырочки не затянулись. Эти поры могут становиться пошире или поуже: либо сами по себе, либо при определённых воздействиях. Каналы могут иметь разное строение, поэтому разные виды каналов имеют разную проницаемость, избирательность и управляемость.
Итак, ионный канал - это интегральный белок, образующий в мембране пору для обмена клетки с окружающей средой ионами K + , Na + , H + , Ca 2+ , Cl - , а также водой, и способный изменять свою проницаемость.
Аквапорины - водные неионные каналы мембраны
В мембране существуют и неионные каналы. Например, аквапорины - это специальные водные каналы , пропускающие через себя воду. Это тоже мембранные каналы, хотя их формально нельзя назвать "ионными каналами".
Пространственная структура канального белка-аквапорина представляет собой цилиндрический канал, по которому движутся молекулы воды. Через него проходит вода и только вода, но не ионы. Аминокислоты в этом белке расположены таким образом, что полярность создаваемого ими электростатического поля переключается в центре канала на обратную. Поэтому молекулы воды, дойдя до середины канала, переворачиваются так, что их дипольные моменты в верхней и нижней части канала оказываются направленными в противоположные стороны. Такое переориентирование предотвращает просачивание через канал заряженных ионов. Аквапорин не пропускает даже ионы гидроксония H3O+ (то есть гидратированные протоны, или ионы водорода), от концентрации которых зависит кислотность среды. При этом клеточный мембранный "водопровод" обладает потрясающей пропускной способностью: он пропускает до миллиарда молекул воды в секунду. Сейчас известно уже около 200 разновидностей белков водных каналов у растений и животных, в том числе 11 - у человека. Благодаря аквапоринам клетки не только регулируют свой объём и внутреннее давление, но и выполняют такие важные функции, как всасывание воды в почках животных и корешках растений.
В настоящее время в молекулярной биологии в основном завершён описательный период в исследовании многообразия катион-транспортирующих ионных каналов в клетках эукариот. Теперь на первый план выходят проблемы познания механизмов регуляции ионных каналов и описание их участия в реакциях живой клетки на различные воздействия и на изменение её микроокружения.
Регулирумый перенос ионов через гидрофильные поры мембраны с помощью управляемых ИК является важнейшим свойством живых клеток, как электровозбудимых, так и невозбудимых.
В связи с этим целесообразно использовать в классификации ионных каналов именно принцип управления их деятельностью. Принцип управления состоянием ионных каналов и был положен в основу предложенной нами () функциональной классификации ионных каналов.
Видео: Ионные каналы в мембране
Строение ИК
ИК состоят из белков сложной структуры (белков-каналоформеров). Схематические изображения ИК приведены ниже, например: .
На рисунке справа показан натриевый канал: вид сверху, с наружной стороны мембраны (Источник: Horn R. (2011). Peering into the spark of life . Nature 475 , 305–306).
Белки ИК имеют определённую конформацию, образующую трансмембранную пору, и "вшиты" в липидный слой мембраны. Канальный белковый комплекс может состоять либо из одной белковой молекулы, либо из нескольких белковых субъединиц, одинаковых или разных по строению. Эти субъединицы могут кодироваться разными генами, синтезироваться на рибосомах по-отдельности и затем собираться в виде целостного канала. В другом случае канал может представлять собой единый полипептид, который в виде петель прошивает мембрану несколько раз. На начало XXI века известно более 400 белков-каналоформеров, для биосинтеза которых используется 1-2% генома человека.
Домены - это отдельные компактно оформленные части канального белка или субъединиц. Сегменты - это части белкка-каналоформера, свёрнутые спирально и прошивающие мембрану. Концевые домены белка-каналоформера (N- и С-терминальные домены) могут торчать из мембраны как наружу, так и внутрь клетки.
Практически все ИК имеют в составе своих субъединиц регуляторные домены , способные связываться с различными управляющими веществами (регуляторными молекулами) и за счёт этого менять состояние или свойства канала. В потенциал-активируемых ИК один из трансмембранных сегментов содержит специальный набор аминокислот с положительными зарядами и работает как сенсор электрического потенциала мембраны. При изменении потенциала такой сенсор меняет состояние канала с открытого на закрытое или наоборот. Таким образом, ИК могут управляться определёнными воздействиями извне, это важное их свойство.
ИК в своём составе могут иметь также вспомогательные субъединицы , выполняющие модуляторные, структурные или стабилизирующие функции. Один класс таких субъединиц - внутриклеточные, расположенные полностью в цитоплазме, а второй - мембранные, т.к. они имеют трансмембранные домены, прошивающие мембрану.
По структуре ИК возможно провести их классификацию, о чём будет сказано ниже.
Свойства ИК
Селективность - это избирательная повышенная проницаемость ИК для определённых ионов. Для других ионов проницаемость понижена. Такая избирательность определяется селективным фильтром - самым узким местом канальной поры. Фильтр, кроме узких размеров, может иметь также локальный электрический заряд. Например, катион-селективные каналы обычно имеют в области своего селективного фильтра отрицательно заряженные остатки аминокислот в составе белковой молекулы, которые притягивают положительные катионы и отталкивают отрицательные анионы, не пропуская их через пору.
Управляемая проницаемость - это способность ИК открываться или закрываться при определённых управляющих воздействиях на канал. Понятно, что закрытый канал имеет пониженную проницаемость, а открытый - повышенную. По этому свойству ИК можно классифицировать в зависимости от способов их открытия: например, потенциал-активируемые, лиганд-активируемые и т.д.
Инактивация - это способность ИК через некоторое время после своего открытия автоматически понижать свою проницаемость даже в том случае, когда открывший их активирующий фактор продолжает действовать.
Быстрая инактивация - это особый процесс со своим особым механизмом, отличающийся от медленного закрытия канала (медленной инактивации). Закрытие (медленная инактивация) канала происходит за счёт процессов, противоположных процессам, обеспечившим его открытие, т.е. за счёт изменения конформации канального белка. А вот, например, у потенциал-активируемых каналов быстрая инактивация происходит с помощью специальной молекулярной "пробки-затычки", напоминающей пробку на цепочке, которую обычно используют в ваннах. Эта пробка представляет собой аминокислотную (полипептидную) петлю с утолщением на конце в виде трёх аминокислот, которым и затыкается внутреннее устье канала со стороны цитоплазмы. Именно поэтому потенциал-зависимые ИК для натрия, обеспечивающие развитие потенциала действия и движение нервного импульса, могут пропускать в клетку ионы натрия только в течение нескольких миллисекунд, а затем они автоматически закрываются своими молекулярными пробками, несмотря на то, что открывающая их деполяризация продолжает действовать. Другим механизмом инактивации ИК может служить модификация дополнительными субъединицами внутриклеточного устья канала.
Блокировка - это способность ИК под действием веществ-блокаторов фиксировать какое-то одно своё состояние и не реагировать на обычные управляющие воздействия. В таком состоянии канал просто перестаёт давать ответы на управляющие воздействия. Блокировку вызывают вещества-блокаторы , которые могут называться антагонистами , блокаторами или литиками .
Антагонисты - это вещества, препятствующие активирующему действию других веществ на ИК. Такие вещества способны хорошо связываться с рецепторным участком ИК, но не способны изменить состояние канала, вызвать его ответную реакцию. Получается блокада рецептора и вместе с ним - блокада ИК. Следует помнить, что антагонисты не обязательно вызывают полную блокаду рецептора и его ИК, они могут действовать более слабо и лишь ингибировать (угнетать) работу канала, но не прекращать её полностью
Агонисты-антагонисты - это вещества, которые обладают слабым стимулирующим влиянием на рецептор, но при этом блокируют действие естественных эндогенных управляющих веществ.
2. Потенциал-управляемые (потенциал-чувствительные, потенциал-зависимые, потенциал-активируемые, voltage-gated). Так, потенциал-управляемые натриевые каналы открываются под действием сдвига электрического потенциала мембраны, превышающего критический уровень деполяризации. Поэтому при достижении определённого порогового уровня деполяризации мембраны они открываются, а при обратном снижении уровня деполяризации - оказываются закрытыми. Но важно знать то, что ещё до обратного снижения уровня деполяризации эти каналы закрываются с внутренней стороны специальными белковыми "пробками" и это происходит автоматически, незасисимо от изменений деполяризации. Вследствие этого потенциал-управляемые натриевые ИК находятся в открытом состоянии всего несколько миллисекунд, а потом закрываются "пробкой", т.е. инактивируются. Окончательно они переходят в закрытое состояние при реполяризации и восстановлении потенциала покоя. Как при химической, так и при фармакологической модификации таких ИК у них сохраняется основной механизм активации и инактивации в ответ на сдвиг мембранного потенциала, что и определяет быстрые изменения катионной проницаемости возбудимых мембран за счёт потенциал-управляемых ИК. Именно такого типа потенциал-управляемые натриевые ИК обеспечивают перемещение нервного импульса по мембране нейрона (смотри: потенциал действия и нервный импульс). Такие потенциал-зависимые натриевые каналы I открываются на уровне КУД, т.е. -55mV, они и формируют потенциал действия и нервный импульс.
Примеры: тетродотоксин-чувствительные натриевые каналы, потенциал-активируемые К-каналы, калиевые Kdr-каналы задержанного выпрямления, кальциевые каналы пресинаптических окончаний аксонов .
На рисунке справа - условная схема работы потенциал-управляемого ИК (кликните на рисунок, чтобы увидеть процесс в динамике).
3. Хемо -управляемые (хемочувствительные, хемозависимые, лиганд-управляемые, лиганд-зависимые, рецептор-активируемые). Они открываются при связывании с рецепторным участком канала специфического лиганда (управляющего вещества: трансмиттера или его миметика). Такие каналы обычно локализованы в химических синапсах на их постсинаптических мембранах и преобразуют химический сигнал, возникающий за счёт пресинаптического высвобождения нейромедиатора, в постсинаптический электрический локальный потенциал. Смотри: локальный потенциал , 3_3 Синапсы , синапсы, медиаторы и модуляторы .
 Примеры: каналы с никотиновыми ацетилхолиновыми рецепторами nAChR), серотониновыми рецепторами (5-HT3), глициновыми, ГАМК-рецепторами (GABAA и GABAC).
Примеры: каналы с никотиновыми ацетилхолиновыми рецепторами nAChR), серотониновыми рецепторами (5-HT3), глициновыми, ГАМК-рецепторами (GABAA и GABAC).
На рисунке справа - лиганд-управляемый ионный канал с никотиновым ацетилхолиновым рецептором (никотин - миметик, ацетилхолин - трансмиттер). Канал состоит из 5 субъединиц и поры в центре. Вверху - поперечный разрез этих субъединиц: a1, a2, b, g, d. Внизу - участки субъединиц, образующих «воротную систему» канала. Представлены аминокислотные последовательности М2 a-спирали в b- и d-. Из 5 субъединиц, образующих пору, изображены лишь 4, а ближайшая к нам удалена, чтобы были видны участки М2, облицовывающие ионный канал, и ворота. Видно, что большая часть молекулы белка выходит за пределы внешней поверхности плазматической мембраны, образуя молекулярные рецепторы к лиганду. Каждая из двух a-субъединица содержит связывающий центр для ацетилхолина, следовательно, с рецептором может связаться 2 молекулы лиганда (нейротрансмиттера или нейромиметика). Ворота, находящиеся в пределах поры, открываются при связывании ацетилхолина с рецепторным участком канала. Отрицательно заряженные остатки глутаминовой и аспарагиновой аминокислот (они выделены синим цветом) имеются в обоих концах М2-спиралей, т.е. с двух сторон поры, благодаря чему предотвращается вход в канал анионов, а катионы Na + и К + при его закрытом состоянии могут быть связаны уже в самом канале.
Видео: Работа хемо-управляемого (лиганд-управляемого) ионного канала
4. Стимул-управляемые (механочувствительные, механосенситивные, стретч-активируемые, stretch-activated, протон-активируемые, температурно-чувствительные).

Они открываются под воздействием специфичного и адекватного для них стимула (раздражителя). Такие каналы обеспечивают сенсорное восприятие и располагаются в мембране сенсорных рецепторов.
Пример: механочувствительные ИК
рецепторных волосковых клеток, обеспечивающих слуховое восприятие; температурно-чувствительные ИК
терморецепторов кожи, обеспечивающие восприятие тепла и холода.
В настоящее время стимул-управляемые механочувствительные ИК обнаружены не только в специализированных механорецепторных структурах, но также и в мембранах бактерий, грибов, растений, позвоночных и беспозвоночных животных. Механочувствительные каналы не только обеспечивают сенсорное восприятие механического раздражения, но также вовлечены в контроль клеточного цикла, регуляцию объёма и роста клеток, секрецию и эндоцитоз.
TRP-каналы в мембране терморецепторов кожи обеспечивают термотрансдукцию, открываясь при различных значениях темпераруры. Они пропускают катионы, особенно ионы кальция.
5. Совместно-управляемые (NMDA-рецепторно-канальный комплекс). Они открываются одновременно как лигандами, так и определённым электрическим потенциалом мембраны. Можно сказать, что у них двойное управление.
Пример: NMDA-рецепторно-канальный комплекс , имеющий сложную систему управления, включающую в себя 8 рецепторных участков-сайтов, с которыми могут связываться различные лиганды.
6. Опосредованно -управляемые (вторично-управляемые, ион-активируемые, ион-зависимые, мессенджер-управляемые, управляемые метаботропными рецепторами). Они открываются и закрываются не под действием прямых внешних сигналов, а вследствие опосредованного воздействия на них внутриклеточных вторичных мессенджеров (ионов кальция Са 2+ , цАМФ, цГМФ, ИФ3, диацилглицерола). Основной механизм такого управления - фосфорилирование ионного канала с внутренней стороны мембраны.
Опосредованное вторичными мессенджерами управление является не прямым, а вторичным. Оно зависит не только от внешнего воздействующего сигнала, но и от наличия, концентрации и активности вторичных мессенджеров. Пусковым сигналом к началу этого процесса может служить воздействие на так называемый метаботропный рецептор , не относящийся к структуре самого управляемого ионного канала и расположенный на мембране где-то отдельно от него. Воздействие на метаботропный рецептор приводит к повышению в клетке концентрации вторичных посредников-мессенджеров. Это ионы Са 2+ , цАМФ, цГМФ, ИФ3, диацилглицерол. Они активируют соответствующие ферменты-протеинкиназы: А-киназы (цАМФ-зависимые), G-киназы (цГМФ-зависимые), В-киназы (кальций-кальмодулин-зависимые) или С-киназы (кальций-фосфолипид-зависимые). В свою очередь, активированные киназы фосфорилируют ИК мембраны изнутри клетки, т.е. присоединяют к ним фосфаты. В результате этого канал может перейди надолго в новое состояние (открытое или, наоборот, закрытое). После срезания фосфатов ферментом фосфатазой канал возвращается к своему прежнему состоянию. В некоторых случаях такой вторичный мессенджер, как G-белок в виде своей активной субъединицы бета-гамма может сам присоединиться к ионному каналу и поменять его состояние. Так, например, могут открываться (активироваться) калиевые каналы при раздражении ацетилхолином мускариновых рецепторов, связанных с G-белком.
Примеры: Са 2+ -активируемые хлорные каналы, кальций-активируемые калиевые каналы, цГМФ-активируемые натриевые каналы палочек сетчатки глаза .
Вот, например, опосредованные ион-управляемые кальций-активируемые хлорные каналы являются одним из основных компонентов системы эпителиальной секреции, сенсорной трансдукции , регулирования нейронной и сердечной возбудимости у животных. В клетках растений кальций-активируемые хлорные каналы ответственны за состояние тургора клетки. При повышении концентрации ионов кальция внутри клетки эти каналы открываются и начинают пропускать ионы хлора.
К опосредованно-управляемым (мессенджер-управляемым) ионным каналам формально можно отнести также все каналы, которые управляются "изнутри" с помощью посредников - вторичных мессенджеров. Такой способ управления чаще всего является дополнительным по отношению к "внешнему управлению", и получается, что к мессенджер-управляемым каналам относится большинство каналов из других групп нашей функциональной классификации. Например, все те ИК, которые могут подвергаться фосфорилированию.
7. Актин-управляемые (актин-регулируемые, actin-regulated, actin-gated channels). Они открываются и закрываются за счёт разборки-сборки примембранных микрофиламентов с участием актин-связывающих белков.
В электроневозбудимых клетках активация и инактивация актин-управляемых потенциал-независимых натриевых каналов контролируется процессами разборки-сборки примембранных микрофиламентов с участием актин-связывающих белков. Актиновые элементы цитоскелета, по-видимому, представляют важнейшую часть потенциал-независимого воротного механизма, управляющего открыванием и закрыванием каналов. Именно сборка микрофиламентов на цитоплазматической стороне мембраны приводит к инактивации таких каналов.
8. Коннексоны (двойные поры). Образуют в мембранах контактирующих клеток сквозные непрерывные каналы через две мембраны сразу в зоне щелевых контактов для взаимного обмена веществами между этими клетками. Через коннексоны передаются электрические сигналы, аминокислоты и небольшие молекулы управляющих веществ: цАМФ, InsP 3 , аденозин, АДФ и АТФ. Они состоят из 6 белковых субъединиц (коннексинов), живущих всего несколько часов. Коннексины - это политопные интегральные мембранные белки 4 раза прошивающие мембрану, имеющие две внеклеточные петли (EL-1 и EL-2), цитоплазматическую петлю (CL) с N-концом (AT) и C-концом (CT), вдающимися в цитоплазму. Через коннексоны соединяется внутренняя среда соседствующих клеток.
Коннексоны являются "неспецифически-управляемыми" каналами. Их состояние регулируется pH, электрическим потенциалом, ионами Са 2+ , фосфорилированием и другими факторами.
Коннексоны найдены практически во всех видах клеток.
9. «Энерго-зависимые транспортёры» (ионные насосы, ионные помпы, ионные обменники, транспортёры). Это особая группа динамичных пор, проводящих ионы через мембрану, которые формально не относятся к ИК. Их деятельность обеспечивается энергией расщепления АТФ. Они представлены мембранными ферментными белками АТФазами, которые активно протаскивают через себя ионы, используя для этого энергию расщепления АТФ, и обеспечивают активный транспорт ионов через мембрану даже против их градиента концентрации.
Примеры: натрий-калиевый насос, протонный насос, кальциевый насос .
Примеры ионных каналов разного типа

Ацетилхолиновый рецептор лиганд-управляемого (хемозависимого) ионного канала
На рисунке слева представлена структурная модель лиганд-управляемого ацетилхолинового ИК.
Последнее обновление: 28/10/2013
Вторая статья из серии «Основы физиологии человека и животных». Речь пойдёт о механизме формирования потенциала действия - основы любого движения.
Возбудимые клетки (которыми являются в той или иной степени все клетки организма животного) в покое имеют избыток отрицательного заряда, формирующий . Если клетка подвергается внешнему раздражению, она переходит в возбуждённое состояние и генерирует другой потенциал - потенциал действия.
Реализует этот процесс система ионных каналов в мембране клетки, регулирующая концентрации электрически заряженных частиц - ионов. Все каналы, независимо от специализации, управляются определёнными силами. Это может быть изменение потенциала на клеточной мембране - в случае потенциал-зависимых каналов, повышение концентрации определённых активных веществ - для лиганд-зависимых или растяжение мембраны - для механически управляемых каналов.
Каналы - это специфические белки, встроенные в мембрану. Каждый тип каналов пропускает определённые ионы. Это система пассивного транспорта: ионы проходят через них благодаря диффузии, а каналы просто контролируют концентрацию проходящих частиц, регулируют для них проницаемость мембраны.
В формировании потенциала действия, как и потенциала покоя, принимают участие главным образом ионы натрия и калия.
Натриевые каналы имеют достаточно простое строение: это белок из трёх разных субъединиц, которые образуют структуру, похожую на пору - то есть трубку с внутренним просветом. Канал может находиться в трёх состояниях: закрытом, открытом и инактивированном (закрыт и невозбудим). Это обеспечивается локализацией отрицательных и положительных зарядов в самом белке; эти заряды притягиваются к противоположным, существующим на мембране, и таким образом канал при изменении состояния мембраны открывается и закрывается. Когда он открыт, ионы натрия могут беспрепятственно проникать через него в клетку по градиенту концентрации. Это очень короткий момент времени - буквально доли миллисекунды.
Калиевые каналы устроены ещё проще: это отдельные субъединицы, имеющие в разрезе трапециевидную форму; они расположены почти вплотную друг к другу, но между ними всегда остаётся зазор. Калиевые каналы не закрываются до конца, в состоянии покоя калий свободно уходит из цитоплазмы (по градиенту концентрации).
И натриевые, и калиевые каналы являются потенциал-зависимыми - они работают в зависимости от изменений электрического потенциала мембраны.
При формировании потенциала действия происходит резкая кратковременная перезарядка мембраны. Это обеспечивается несколькими последовательными процессами.
Сначала под воздействием внешнего раздражения (например, электрического тока) мембрана деполяризуется - то есть заряды с разных её сторон меняются на противоположные (внутри клетки заряд переходит в положительный, снаружи - в отрицательный). Это является сигналом к открытию натриевых каналов, которых на поверхности одной мембраны огромное число - может быть до 12 тысяч. Момент, в который начинают открываться каналы, носит название критического уровня деполяризации. Ток, который даёт эту критическую деполяризацию, назван пороговым.
Интересно, что повышение силы тока после достижения пороговой величины не меняет характеристик получающегося в итоге потенциала действия. Значение для открытия каналов имеет не амплитуда тока, а полученное мембраной количество энергии - «количество электричества». Эта закономерность получила название «всё или ничего» - либо есть полноценный ответ на раздражение при его величине от пороговой и выше, либо ответа нет вообще, если раздражение пороговой величины не достигло. При этом значение пороговой величины определяется длительностью подаваемого раздражения.
Действителен этот закон, правда, только в рамках отдельной клетки. Если брать, например, нерв, составленный большим количеством разных аксонов, амплитуда тоже будет иметь значение, потому что ответ на раздражение мы увидим только тогда, когда каналы активируются во всех клетках - то есть при большем суммарном значении порогового тока.
После открытия каналов натрий начинает поступать в клетку, и его ток значительно превышает ток выходящего по градиенту калия. Это значит, что проницаемость мембраны для натрия становится больше, чем для калия. В определённый момент открываются почти все натриевые каналы. Это происходит лавинообразно: от той точки, в которую пришёл стимул, в обе стороны. Таким образом, концентрация натрия в клетке резко повышается.
После этого концентрации ионов должны вернуться к исходным. Это обеспечивает такое общее свойство каналов, как рефрактерность: канал, который сработал, некоторое время после этого неактивен и не может возбудиться под действием раздражающего стимула.
Натриевые каналы в момент максимального ответа на раздражение становятся рефрактерны, проницаемость для натрия резко падает. Калиевые каналы, напротив, начинают активно работать, и ток калия из клетки возрастает. Таким образом из клетки уходит избыток положительно заряженных ионов и восстанавливается изначальный потенциал покоя. Этот период времени, пока не восстановятся натриевые каналы и исходный потенциал (это может занимать около миллисекунды), клетка не способна возбудиться.
Поскольку способность клеток к возбуждению обеспечивает работу организма как целого и возможность центрального контроля всех клеток организма, яды, блокирующие каналы, являются одними из самых опасных для человека и многих животных.
Один из самых страшных блокаторов каналов - тетродотоксин, вещество, вырабатываемое рыбой фугу. Для него значение LD50 (50% Level of Death - доза, от которой умрут 50 человек из ста) равно 10 миллиграмм на килограмм веса, то есть примерно в тысячу раз меньше, чем для цианида. Его молекулы связываются прочной связью с белком натриевого канала, когда он в закрытом состоянии, и полностью блокируют возможность возникновения потенциала действия. Похожие токсины вырабатывают некоторые водоросли. Яд скорпиона, напротив, держит все каналы в постоянно открытом состоянии.
Ну ладно скорпион, а вот зачем такое страшное оружие водорослям - загадка.
Есть что сказать? Оставть комментарий!.
Сквозь мембрану. Такие комплексы представляют собой набор идентичных или гомологичных белков, плотно упакованных в липидном бислое мембраны вокруг водной поры. Каналы расположены в плазмалемме и некоторых внутренних мембранах клетки.
Через ионные каналы проходят ионы Na + (натрия), K + (калия), Cl − (хлора) и Ca 2 + (кальция). Из-за открывания и закрывания ионных каналов меняется концентрация ионов по разные стороны мембраны и происходит сдвиг мембранного потенциала.
Канальные белки состоят из субъединиц, образующих структуру со сложной пространственной конфигурацией, в которой кроме поры обычно имеются молекулярные системы открытия, закрытия, избирательности, инактивации, рецепции и регуляции. Ионные каналы могут иметь несколько участков (сайтов) для связывания с управляющими веществами.
Энциклопедичный YouTube
1 / 3
Калий-натриевый насос.flv
2016_03_12 Роль ионных каналов и ионных насосов в регуляции мышечного сокращения - Рубцов А.М.
ЛЕКЦИЯ НОБЕЛЕВСКОГО ЛАУРЕАТА ПО ФИЗИОЛОГИИ И МЕДИЦИНЕ ЭРВИНА НЕЕРА В КАЗНМУ
Субтитры
Типы ионных каналов
Классификация ионных каналов проводится по различным параметрам и поэтому единой унифицированной классификации для них пока не существует.
Так, возможна классификация по структуре (строению) и происхождению от однотипных генов .
По этому принципу, например, выделяют три семейства лиганд-активируемых ионных каналов :
- с пуриновыми рецепторами (АТФ-активируемые);
- с никотиновыми АХ-рецепторами , ГАМК -, глицин - и серотонин -рецепторами ;
- с глутаматными рецепторами .
При этом в одно и то же семейство попадают ионные каналы с разной ионной селективностью, а также с рецепторами к разным лигандам . Но зато образующие эти каналы белки имеют большое сходство в строении и происхождении.
Ионные каналы также можно классифицировать по селективности в зависимости от проходящих через них ионов : натриевые, калиевые, кальциевые, хлорные , протонные (водородные).
Согласно функциональной классификации , ионные каналы группируются по способам управления их состоянием на следующие виды:
- Неуправляемые (независимые).
- Потенциал -управляемые (потенциал-чувствительные, потенциал-зависимые, voltage-gated).
- Лиганд-управляемые (хемо-управляемые, хемочувствительные, хемозависимые, лиганд-зависимые, рецептор -активируемые).
- Опосредованно-управляемые (вторично-управляемые, ион-активируемые, ион-зависимые, мессенджер -управляемые, управляемые метаботропными рецепторами).
- Совместно-управляемые (NMDA-рецепторно-канальный комплекс). Они открываются одновременно как лигандами, так и определённым электрическим потенциалом мембраны . Можно сказать, что у них двойное управление. Пример: NMDA-рецепторно-канальный комплекс, имеющий сложную систему управления, включающую в себя 8 рецепторных участков-сайтов, с которыми могут связываться различные лиганды.
- Стимул -управляемые (механочувствительные, механосенситивные, активируемые растяжением (stretch) липидного бислоя, протон -активируемые, температурно-чувствительные).
- Актин -управляемые (актин-регулируемые, actin-regulated, actin-gated channels).
- Коннексоны (двойные поры).
Наиболее часто встречаются два типа каналов: ионные каналы с лиганд-зависимыми воротами (находятся, в частности, в постсинаптической мембране нервно-мышечных соединений) и ионные каналы с потенциал-зависимыми воротами. Лиганд-зависимые каналы превращают химические сигналы, приходящие к клетке, в электрические; они необходимы, в частности, для работы химических синапсов . Потенциал-зависимые каналы нужны для распространения потенциала действия .
Работа ионных каналов
Неуправляемые (независимые) ионные каналы
Эти каналы обычно находятся в открытом состоянии и постоянно пропускают через себя ионы за счёт диффузии по градиенту их концентрации и/или по электрическому градиенту зарядов по обе стороны мембраны. Некоторые неуправляемые каналы различают вещества и пропускают через себя по градиенту концентрации все молекулы меньше определённой величины, их называют «неселективные каналы» или «поры». Существуют также «селективные каналы», которые благодаря своему диаметру и строению внутренней поверхности переносят только определённые ионы. Примеры: калиевые каналы , участвующие в формировании мембранного потенциала покоя, хлоридные каналы , эпителиальные натриевые каналы , анионные каналы эритроцитов.
Потенциал-зависимые ионные каналы
Эти каналы отвечают за распространение потенциала действия, они открываются и закрываются в ответ на изменение мембранного потенциала . Например, натриевые каналы. Если мембранный потенциал поддерживается на уровне потенциала покоя , натриевые каналы закрыты и натриевый ток отсутствует. Если мембранный потенциал сдвигается в положительную сторону, то натриевые каналы откроются, и в клетку начнут входить ионы натрия по градиенту концентрации . Через 0,5 мс после установления нового значения мембранного потенциала, этот натриевый ток достигнет максимума. А еще через несколько миллисекунд падает почти до 12. Во время покоя мембранного потенциала , внутриклеточная концентрация ионов натрия 12 ммоль/литр, а внеклеточная концентрация 145 ммоль/литр. Это значит, что каналы через некоторое время закрываются вследствие инактивации, даже если клеточная мембрана остается деполяризованной. Но закрывшись, они отличаются от состояния, в котором находились до открытия, теперь они не могут открываться в ответ на деполяризацию мембраны, то есть они инактивированны. В таком состоянии они останутся до тех пор, пока мембранный потенциал не вернется к исходному значению и не пройдет восстановительный период, занимающий несколько миллисекунд.
Лиганд-зависимые ионные каналы
Эти каналы открываются, когда
Селективность - это избирательно повышенная проницаемость ионного канала для определённых ионов и пониженная для других. Такая избирательность определяется селективным фильтром - самым узким местом канальной поры. Фильтр, кроме узких размеров, может иметь также локальный электрический заряд.
Управляемая проницаемость - это способность открываться или закрываться при определённых управляющих воздействиях на канал.
Инактивация - это способность ионного канала через некоторое время после своего открытия автоматически понижать свою проницаемость даже в том случае, когда открывший их активирующий фактор продолжает действовать.
Блокировка - это способность ионного канала под действием веществ-блокаторов фиксировать какое-то одно своё состояние и не реагировать на обычные управляющие воздействия. Блокировку вызывают вещества-блокаторы, которые могут называться антагонистами, блокаторами или литиками.
Родерика Маккинона (Roderick MacKinnon) . Работа содержит проволочный каркас, удерживающий выдутый из желтого стекла объект, который репрезентирует основную полость канальной структуры.
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ И ИОННЫЕ КАНАЛЫ
Биологические мембраны – это функционально активные структуры клеток, ограничивающие цитоплазму и большинство внутриклеточных структур; образуют единую внутриклеточную систему канальцев, складок и замкнутых полостей.
Структурная основа мембраны – двойной слой фосфолипидов, в который встроены мембранные белки. Толщина клеточных мембран 6-12 нм. Молекулы липидов амфотерны. Своими гидрофильными частями они обращены в сторону водной среды (межклеточная жидкость и цитоплазма), гидрофобные части молекул направлены внутрь фосфолипидного бислоя. Такая структура идеально подходит для раздела внеклеточной и внутриклеточной фаз.
Белки, интегрированные в двойной слой фосфолипидов своими полярными участками, образуют гидрофильную поверхность в водной фазе. Они выполняют различные функции: рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул.
Большинство наших знаний об устройстве ионного канала, является результатом функциональной реконструкции. Каждый канал имеет устье, селективный фильтр, ворота и механизм управления воротами.
Часть каналов являются электроуправляемыми, т.е. управляются за счет разности потенциалов на мембране (потенциал-зависимые ионные каналы). Для этого рядом с каналом имеется электрический сенсор, который в зависимости от величины мембранного потенциала либо открывает ворота каналов, либо держит их закрытыми.
Второй вариант ионных каналов – рецептороуправляемые. Ворота управляются за счет рецептора, расположенного на поверхности мембраны (открываются при взаимодействии медиатора с рецептором). В некоторых рецептороуправляемых каналах между рецептором и воротным механизмом имеется промежуточная стадия (посредник типа цАМФ, протеинкиназы и т.д.)
Ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость.
Селективность – или избирательность канала обеспечивается его особой белковой структурой, геометрией канала.
Например, диаметр иона натрия – 0,19 нм, вместе с гидратной оболочкой он становится около 0,3 нм. Устье натриевого канала 0,3 – 0,5 нм. Чтобы пройти через канал (особенно через селективный фильтр), ион натрия или другой ион должен освободиться от гидратной оболочки и только в «голом» виде может пройти через канал. Слишком большой ион не может войти в устье, слишком маленький не способен отдать гидратную оболочку в селективном фильтре, поэтому не может выскочить из канала.
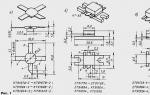
Натриевые каналы (рис. 6) имеют ворота 2-х типов – активационные (m-ворота) и инактивационные (h-ворота). В условиях покоя активационные ворота закрыты, но готовы в любую минуту открыться, а инактивационные – открыты. При снижении МП (деполяризация до 60 мВ) активационные ворота открываются и впускают ионы натрия в клетку, но вскоре начинают закрываться инактивационные ворота (происходит инактивация натриевых каналов). Некоторое время спустя закрываются активационные ворота, открываются инактивационные, и канал готов к новому циклу. Канал блокируется тетродотоксином, местными анестетиками (новокаин и др.).
Рис. 6. Работа натриевых каналов и «воротных» механизмов.
А – в покое m-ворота закрыты; Б – при возбуждении m-ворота открыты; В – закрытие h-ворот (инактивация) при деполяризации.
Калиевые каналы тоже достаточно селективны – в основном пропускают ионы калия. Блокируются тетраэтиламмонием. Процессы инактивации у них выражены слабо. Зато имеются особые калиевые каналы, активирующиеся при повышении внутриклеточной концентрации кальция и деполяризации клеточной мембраны. Активация калий-калльцийзависимых каналов ускоряет реполяризацию (восстановление МП покоя).
Кальциевые каналы. Входящий кальциевый ток недостаточно велик, чтобы нормально деполяризовать клеточную мембрану. Кальций выступает в роли вторичного
посредника (мессенджера). Активация кальциевых каналов обеспечивается деполяризацией клеточной мембраны, напр., входящим натриевым током. Инактивация кальциевых каналов происходит при повышении внутриклеточной концентрации свободного кальция. Однако белки цитоплазмы связывают кальций, что позволяет некоторое время поддерживать стабильную величину кальциевого тока, хотя и на низком уровне; при этом натриевый ток полностью подавляется. Блокируются кальциевые каналы ионами марганца, никеля, кадмия (2-хвалентные ионы), а также лекарственными веществами (верапамил).
Различают пассивный (без затрат энергии) и активный (энергозависимый) транспорт ионов через мембраны.
Пассивный идет за счет простой и облегченной диффузии.
Простая диффузия идет в соответствии с законом Фика – по химическому, электрохимическому или осмотическому градиенту. Напр., в клетке натрия 14 ммоль, а в среде 140 ммоль, в этом случае пассивный поток должен быть направлен в клетку.
Для пассивной простой диффузии вещество должно быть жирорастворимым. Гидрофильные вещества в фосфолипидном бислое мембраны пройти не могут.
Облегченная диффузия происходит или при наличии специализированных каналов или с участием переносчиков, которые специфически связываются с переносимой молекулой, а затем способствуют ее переносу по градиенту концентрации.
Активный транспорт ионов насосами клеточных мембран обеспечивает поддержание ионных градиентов по обе стороны мембраны. Энергия затрачивается на перенос данного вещества против градиента его концентрации.
Доказано участие в активном транспорте ионов специализированных ферментных систем – АТФ-аз, которые осуществляют гидролиз АТФ. Различают:
Натрий–калиевая–АТФ–аза («натриевый насос») обнаружена в мембранах клеток всех животных, растений и микроорганизмов. Это мембранный белок, имеющий два центра связывания ионов. Один из них (натриевый) расположен на внутренней поверхности клеточной мембраны, второй (калиевый) – на ее внешней поверхности. Специфическим ингибитором фермента является сердечный гликозид – строфантин (уабаин), блокирующий работу натриевого насоса. Гидролиз одной молекулы АТФ сопровождается выведением из клетки трех ионов натрия и закачиванием в клетку двух ионов калия. При увеличении количества ионов калия во внеклеточной среде или ионов натрия внутри клетки работа насоса усиливается.
Кальциевая–АТФ-аза («кальциевый насос») наиболее широко распространена в мембранах саркоплазматического ретикулума мышечных клеток.
Протонная–АТФ-аза («протонный насос») – в мембранах митохондрий.